Hormonas y Ejercicio
José L Ferretti0
Resumen
Palabras Clave: sistema endocrino, sistemas reguladores, ejercicio, insulina, hipotálamo, factores reguladores
INTRODUCCIÓN
Se ha visto que todas las manifestaciones vitales de las estructuras vivientes (y con ellas toda expresión de movimiento como las que atañen al ejercicio físico) constituyen el resultado de, al menos, una reacción química, sin excepción (1). Esto puede extrañar al no iniciado en el estudio de los fenómenos biológicos, por cuanto son perfectamente conocidas algunas manifestaciones vitales extraordinariamente rápidas (batir las alas de los insectos, ejecución de un trino de un pianista, etc.) para las cuales resulta casi increíble concebir la producción de fenómenos bioquímicos paralelos.
La credibilidad se restringe más aún si se tiene en cuenta que las reacciones químicas en cuestión son, en todos los casos, de naturaleza orgánica, y que las reacciones orgánicas cursan, en condiciones naturales con extraordinaria lentitud.
El secreto de esa velocidad, como muchos sabemos, consiste en la participación de aceleradores (catalizadores) moleculares llamados enzimas, que unen a una especificidad más o menos alta de acción (no se equivocan de reacción), una eficiencia tremenda en su tarea de buscar, encontrar, y enlazar a las moléculas participantes en las reacciones que catalizan. Aún así, sin embargo, llama poderosamente la atención que solo la velocidad y la especificidad de acción están resueltas con recursos moleculares en los seres vivos, sino que también lo están la regulación de la intensidad y la determinación de la intensidad, y la determinación de la oportunidad (de comienzo, de aceleración, de mantenimiento, de desaceleración, de detención) de las reacciones (relaciones de “orden” (2)), y que estas relaciones de orden están coordinadas con tan maravillosa precisión como para hacer pensar a los eventuales observadores de cada proceso que el mismo entraña algún “objetivo funcional”, o, más esotérica o metafísicamente hablando, alguna finalidad.
Ya hemos descartado a priori esta clase de interpretaciones reñidas con los cánones del pensamiento científico (2, 3). Llega ahora el momento de ofrecer algunas de las explicaciones biológicamente razonables para la producción de los fenómenos determinantes de esas relaciones de orden. Obviamente, en ningún caso podremos apartarnos, según lo que hemos venido comentando, de la participación de alguna clase de enzima en el proceso en cuestión. El trabajo catalizador de las enzimas puede acelerarse o retardarse mediante una gran variedad de fenómenos: desde el estímulo o la inhibición de su producción por los ribosomas citoplasmáticos mediante efectos sobre los ácidos nucleicos (genes) respectivos, hasta la variación de la temperatura, del ph, de la concentración de las sustancias necesarias como complementos moleculares (cofactores) de las enzimas, o simplemente de las sustancias capaces de acelerar o retardar el trabajo enzimátco mediante simples interacciones intermoleculares que modifican la afinidad de las enzimas con su sustrato (inhibidores o activadores enzimáticos).
Lógicamente, la selección natural, vehículo de la Evolución, ha favorecido el desarrollo de las estructuras vivientes primitivas (unicelulares) capaces de reaccionar a la variación agresiva de su medio ambiente (externo) con la producción de cambios que, directa o indirectamente, modificarán adecuadamente la actividad de las enzimas implicadas en la producción de las manifestaciones biológicas tendientes a neutralizar (o aún a trasformar en útiles) los efectos de los cambios ambientales. No es de extrañar, entonces, que las especies unicelulares que se han mantenido evolucionando hasta nuestros días, tras tres mil quinientos millones de años de interacción y prueba de “ensayo y error” versus el ambiente, posean mecanismos (enzimáticos) adaptatorios casi perfectos para resolver una enorme gama de situaciones, derivadas tanto de la agresión como de la solicitación positiva (“motivación”) a partir de su ambiente.
La solución evolutiva recorrió caminos mucho más tortuosos, sin embargo, cuando el mismo problema debió encararse para los organismos pluricelulares hace mil millones de años. En efecto, en los pluricelulares, conviene obviamente que los cambios adaptativos sean adoptados por todas las células de una misma clase.
Cuando las células “aprendieron” a vivir de a dos, o en grupos pequeños, en los que prácticamente todas podían contactar unas con otras, con mínimas diferencias metabólicas entre ellas, la coordinación temporal de la producción de cambios adaptativos representó simplemente un problema de comunicación intercelular “membrana por medio” de solución comparativamente fácil: bastaba la posibilidad del paso de ciertas moléculas de una célula a su vecina para “homologar situaciones”.
Cuando la incorporación de un número mayor de células por la estructura obligó a la pérdida de contacto “por simple deformación natural” de las células vecinas, comenzaron a desarrollarse prolongaciones especiales de algunas de ellas, que les permitían mantenerse en comunicación con su (s) “compañera (s) de trabajo” mediante el mismo mecanismo de pasaje de sustancias (ya podemos llamarlas “mediadores químicos” de información) a través de los puntos contactantes de sus membranas. Esta forma de adaptación del sistema de comunicación, caracterizada por mantener totalmente la especificidad del contacto (jamás puede “equivocarse de compañera”) a pesar de la distancia (que a veces llegó a ser muy grande) dio lugar a lo que hoy conocemos como Sistema Nervioso. Su perfeccionamiento, gracias al desarrollo de la capacidad de conducción de la información en forma de cambios alternantes de la polaridad de las membranas de las prolongaciones (ahora dendritas y axones), cuyo arribo a los puntos de contacto célula/célula induce a la liberación y el paso de los mediadores a la otra unidad de trabajo, incrementó además enormemente la velocidad de comunicación.
Naturalmente, no todos los problemas de comunicación interna de los pluricelulares pudieron vencerse de esta misma manera. Al contrario, en la mayoría de los casos, las células se iban separando durante el desarrollo, adquiriendo al mismo tiempo determinadas especializaciones funcionales que las distinguían de sus ex –vecinas. Este proceso determinó el desarrollo de una “reparación funcional del trabajo” que ofreció enormes ventajas competitivas (es decir, selectivo-evolutivas) a las especies que lo conformaron, pero que sólo pudieron efectivizarse si, al mismo tiempo que la diferenciación se producía, tenía también lugar un mecanismo mantenedor (o generador, si antes no lo había) de la necesaria comunicación tejido a tejido (ya más que célula a célula) para proveer la adecuada coordinación de los diferentes trabajos de cada subestructura especializada podía ejecutar, sea con exclusividad o, al menos, más perfectamente que otras.
En algunos casos menos complicados, en que las células necesitadas de comunicación permanecían más o menos cercanas y no se especializaban en forma distintiva (es decir, seguían siendo de la misma clase), o bien adquirían diferenciación unas de otras, pero no demasiadas, el problema se dirimió mediante la liberación, por algunas de ellas, de sustancias que pasaban al medio acuoso intercelular cercano y eran captadas por las demás, sean de su misma clase (fenómeno autocrino) o especializadas en otras funciones (fenómeno paracrino). Cuando la distancia (y obviamente las diferencias funcionales) entre las células emisoras y receptoras de “información molecular” se hizo demasiado grande como para ser salvada por simple difusión intratisular, la comunicación solo fue posible aprovechando el desarrollo simultáneo (organismos complejos) del sistema circulatorio. Este recurso permitió expandir prácticamente al infinito las posibilidades de comunicación entre los rincones más remotos del organismo, entre células, tejidos y aún órganos de distintas clases, aunque basándose estructuralmente siempre en mecanismos integrados a nivel celular de complejidad (2). Sustancias elaboradas en cualquier sitio del organismo podían, efectivamente, alcanzar cualquier otro sitio para provocar allí, por simple presencia o por variación de su concentración en el ambiente interno local, cambios de cualquier naturaleza. En principio, lo de “cualquiera” es válido; pero es fácil caer en la cuenta que, si el resultado óptimo del desarrollo de estos mecanismos de comunicación biológica es precisamente la provisión y el perfeccionamiento de las “relaciones de orden”, no es precisamente lo ideal que pueda ocurrir cualquier cosa en cualquier parte. Fue preciso, para evitar este eventual caos informativo, la ocurrencia de por lo menos dos fenómenos, respectivamente, en las células emisoras de mensajes y en las receptoras:
a) Una paulatina diversificación entre las sustancias mediadores de la información: esto implicó un aumento de la complejidad de las moléculas posibles de utilizarse como vectores de mensajes por las células emisoras, porque a mayor complejidad es fácil conseguir también mayor posibilidad de diversidad en cualquier molécula.
b) El desarrollo de la capacidad de producir moléculas específicas por parte de las células receptoras, para “identificar” bioquímicamente a las moléculas “mensajeras”. Estas moléculas especiales se denominan genéricamente “receptores”. Pueden ubicarse tanto sobre las membranas celulares cono en el seno del citoplasma, y poseen una conformación espacial específica, que les permite acoplarse con las moléculas “mensajeras” por simple afinidad intermolecular derivada de la distribución de cargas eléctricas sobre su superficie (la cual deviene a su vez de la conformación molecular espacial).
De la interacción (acople) entre las moléculas mensajeras y receptoras resulta un complejo molecular que a su vez, es capaz de ejercer una actividad catalítica sobre diversos procesos bioquímicos celulares.
Este sistema particular de “transmisión de información biológica a distancia” se denomina endocrino, y las sustancias encargadas de transportar la información se las conoce como hormonas. Las hormonas, pues, no son otra cosa que moléculas producidas dentro de un organismo pluricelular complejo generalmente en cantidades muy pequeñas (porque actúan “encontrando específicamente a sus receptores” y no por acción masiva de su simple concentración), que ejercen importantes acciones biológicas a distancia de donde fueron producidas y liberadas, interactuando con otras moléculas (receptores) propias de las células a las que debe llegar la información (“células blanco” o “target cells”). Generalmente, los efectos resultantes de la acción de las hormonas conllevan a su vez cambios biológicos que pueden actuar como inductores o inhibidores de la producción de la hormona original, de modo que esta puede considerarse a su vez regulada por un mecanismo retroalimentado (4) (Figura 1).
La distancia y la diversidad de posibles “células blanco” obligan a una especificidad muy grande en la conformación espacial (estructura) de la molécula hormonal y de sus receptores, que por esta razón deben ser de una complejidad bastante más alta que la de un simple mediador de información “por contigüidad”, como los que operan en el Sistema Nervioso.

Figura 1
La complejidad molecular de las hormonas es especialmente importante por esta razón, al punto que se describen dos grandes grupos de hormonas respecto de la forma de interactuar con sus receptores:
a) Las hormonas de tipo esteroide, de naturaleza lípida, que por esta misma razón pueden atravesar fácilmente las membranas celulares y acoplarse con receptores citoplasmáticos. Estas hormonas ejercen generalmente sus efectos mediante la interacción de los complejos hormona-receptor con el ADN del núcleo celular, es decir, mediante la inducción o la represión del trabajo de un gen de la célula “blanco” (efecto generalmente lento; Figura 2a).
b) Las hormonas del tipo peptídico, que, insolubles en el material lipídico de las moléculas celulares, actúan acoplándose a moléculas receptoras alojadas en la misma membrana, de modo que la formación del complejo hormona-receptor determina la producción de efectos catalíticos del lado citoplasmático, como resultado de los cuales se forman nuevas moléculas intracelulares, llamadas “segundos mensajeros”, que a su vez actúan estimulando o inhibiendo determinadas reacciones químicas en el citoplasma y produciendo de esta forma el efecto final de la hormona (mecanismo generalmente rápido); (Figura 2b).
La complejidad de los mecanismos de producción de los fenómenos hormonales va de la mano con la necesidad de su especificidad de acción, que a veces debe ser muy alta; y expone también una parte importante de la gama de recursos con los que los organismos complejos cuentan para cumplimentar las mentadas “relaciones de orden” al son del trabajo de simples moléculas.
Todos los mecanismos descriptos de comunicación intercelular tienen importancia para la producción de las manifestaciones vitales asociadas con el movimiento implícito en el ejercicio físico. Los procesos auto y parácrinos intervienen en la inducción del desarrollo de las estructuras del aparato osteomuscular, garantizando la diferenciación y especialización local de sus diversas estructuras. La coordinación nerviosa es primordial para la transmisión y coordinación del impulso motor. Y los fenómenos endocrinos son responsables: a)de la interrelación que existe entre el trabajo muscular y el de los tejidos proveedores de nutrientes (adiposo y hepático); b) de la repercusión que el ejercicio provoca sobre gran cantidad de órganos, aparatos y sistemas.
A continuación trataremos la forma de cómo el sistema endocrino responde al trabajo muscular, aspecto del metabolismo indispensable para la comprensión actual de la fenomenología que estudia la Fisiología del Ejercicio.

Figura 2
CONTROL DEL METABOLISMO HIDROCARBONADO DURANTE EL EJERCICIO
1. Consideraciones Generales
En un sujeto en reposo, el metabolismo está sujeto en gran parte a un control homeostático a cargo de sistemas regulatorios retroalimentados, la mayoría de ellos de carácter negativo (tendientes a mantener los valores de determinadas variables biológicas dentro de márgenes “normales” de variación (4)).
En el caso del metabolismo hidrocarbonato, que comprende procesos esenciales para la provisión energética del músculo durante el ejercicio físico, la velocidad y en sentido de las reacciones químicas implicadas, catalizadas siempre por enzimas específicas, se encuentran bajo el control de una gran variedad de mediadores químicos (mensajeros moleculares de información) que actúan entre células de distinta estirpe.
Los mediadores químicos en cuestión son invariablemente hormonas (2), cuya interacción con los receptores de las membranas del citoplasma de las “células blanco” termina siempre afectando directa o indirectamente alguna clase de cambio en el trabajo metabólico celular, tendiente generalmente a adecuarlo a las distintas condiciones establecidas por la tasa de actividad muscular circunstancial del organismos.
El principal problema a resolver dentro de este contexto durante el ejercicio, es la provisión a los músculos del combustible adecuado en calidad, cantidad y oportunidad para la clase de trabajo que se está realizando. Esta tarea se lleva a cabo automática e imperceptiblemente para cada individuo gracias a las interrelaciones establecidas entre las variables biológicas correspondientes a los mediadores hormonales, que configuran parte de la organización de su vida vegetativa.
Tanto las estructuras orgánicas que aportan combustibles (fundamentalmente el hígado, que aporta glucosa, y el tejido adiposo, que proporciona ácidos grasos libres- AGL-) como las que los consumen (músculo, cerebro, riñones, etc.) lo hacen utilizando a la sangre circulante como vehículo de transporte. Por consiguiente, la concentración de cada clase de combustible en la sangre, que varía a cada instante de acuerdo con las fluctuaciones de las correspondientes tasas de aporte y de consumo, constituye un indicador adecuado de las respectivas velocidades de provisión y de utilización. También la circulación constituye el medio adecuado para encauzar a las moléculas hormonales, permitiéndoles alcanzar a las respectivas células-blanco para ejercer sus efectos a distancia de donde son producidas.
No debe extrañar, en consecuencia, que en la generalidad de los casos, las interrelaciones entre las variables biológicas de cuya interacción resulta el control del aprovisionamiento de combustible para el trabajo muscular de todo el cuerpo reconozcan dos clases fundamentales de mensajes:
a) La variación de la concentración sanguínea de un combustible, originada por cambios en la tasa de trabajo muscular voluntario o en el aporte por las distintas fuentes, es detectada por alguna estructura (glándula endocrina) productora de un mediador químico (hormona) capaz de ser reconocido por los receptores de otra estructura diferente, de alguna manera, tiene que ver con el aporte y/o con el consumo de ese combustible. El resultado de este paso del proceso regulatorio es una variación adecuada (aumento o disminución según el caso) de la producción del mediador químico, que se vuelca a la circulación, haciendo variar obviamente su propia concentración sanguínea:
Concentración de combustibles >>>>> Glándula Endocrina >>>>> Hormona
b) La variación de la concentración sanguínea de la hormona es detectada específicamente (sin posibilidad de error, gracias a la participación de receptores molecularmente específicos) por la segunda estructura (que por esta razón se denomina “estructura blanco”). El resultado de este segundo paso del mecanismo regulatorio es una variación (aumento o disminución), igualmente adecuada para el caso, de la liberación o de la captación del combustible a/o desde la sangre circulante, cuyo resultado, en términos de concentración sanguínea del combustible, es generalmente opuesto al de la variación inicial considerada en (a);
Hormona >>>>> Estructura “blanco” >>>>> Concentración de combustibles
La integración de los pasos (a) y (b) en un ciclo único, gracias a la participación de la circulación sanguínea, constituye un sistema regulatorio elemental del tipo retroacoplado (retroalimentado, o de feed-back (5)), que puede esquematizarse en forma sobresimplificada tal como en la Figura 3.
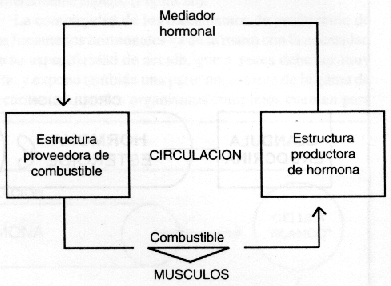
Figura 3
En el caso particular del metabolismo hidrocarbonato, el principal combustible circulante es, por lejos, la glucosa (tan así es, que se puede, en la práctica, hacer abstracción de la participación de otros glúcidos simples, como la galactosa o la fructosa, como aporte energético para el ejercicio). La principal estructura proveedora, en ausencia del trabajo intestinal absortito de los alimentos, es el hígado; y las estructuras productoras de hormonas en relación con las fluctuaciones sanguíneas del combustible son varias glándulas endocrinas, cada una de las cuales participan de un modo distinto en el sistema.
El funcionamiento general del mecanismo sólo puede esquematizarse en forma simple si se se considera que su comportamiento, en cuanto a un sistema regulatorio negativamente retroalimentado, “apunta” (competitivamente, en términos evolutivos) al mantenimiento de una cierta constancia de los valores de una sola de las variables sanguíneas: la concentración de glucosa en la sangre, o glicemia. No siendo por la concentración de ninguna de las hormonas reguladoras, cuya fluctuación descontrolada (especialmente hacía abajo) podría acarrear inconvenientes muy serios al funcionamiento de muchas estructuras del organismo, especialmente al cerebro, y entendiendo de esta forma, el sistema puede representarse gráficamente teniendo en cuenta la existencia de una “variable regulada” (la glicemia) cuya “perturbación” (por el trabajo muscular, en este caso) resulta neutralizada por la producción (por parte de las glándulas endocrinas) de varios “reguladores” hormonales (4):
Perturbación >>>>> Variable regulada <<<<< Sistema regulador
Trabajo muscular >>>>> Glicemia <<<<< Glándulas endocrinas
Las principales glándulas endocrinas implicadas en estos mecanismos pueden clasificarse en dos grupos, de acuerdo con el signo de su respuesta (aumento o disminución de la producción de sus respectivas hormonas) frente a una variación dada de la glicemia. Por un lado, el páncreas endocrino productor de insulina (células beta de los islotes de Langerhans), hormona hipoglicemiante, cuya secreción aumenta o disminuye en forma directamente proporcional al aumento o a la disminución de la glicemia; y por otro, las glándulas productoras de las hormonas llamadas “antiinsulínicas” o “protectoras del ayuno”, de efecto hiperglucemiante (células eosinófilas de la adenohipófisis, capa fasciculada de la corteza suprarrenal, el sistema adrenérgico, células alfa de los islotes pancreáticos, tiroides), cuya secreción aumenta o disminuye en proporción inversa con als variaciones glicéricas:

2. Mecanismos elementales de acción de las hormonas reguladores
Con la excepción de las hormonas producidas por la corteza suprarrenal, que poseen estructura molecular esteroide (lipídica), las restantes hormonas (polipeptídicas o de molécula simple no lipídica) ejercen su acción sobre las células “blanco” mediante la activación de mecanismos de membrana que involucran la producción y participación del AMP cíclico (2), un “segundo mensajero” de la información cuyo papel, generalmente limitado a la activación o la inhibición de una enzima clave para el funcionamiento de una determinada cadena de reacciones metabólicas, puede asimilarse al de una “hormona intercelular”.
La insulina, hormona peptídica, incrementa la captación de la glucoas sanguínea por el hígado y por todos los otros tejidos consumidores (salvo el cerebro y los glóbulos rojos), mediante algunos de los siguientes mecanismos, facilitando su transporte a través de las membranas celulares aprovechando la participación de las moléculas transportadoras (“carriers”):
a) Promoviendo su rápida fosforilación intracelular a glucosa-6-fosfato (que ya no puede volver a salir de la célula) por la enzima hexoquinasa, lo que permite su ulterior metabolización (evitando prácticamente la presencia de glucosa como tal en el citoplasma);
b) Inhibiendo la formación de la glucosa a partir de glucosa-6-fosfato por la enzima glucosa-6-fosfatasa, y favoreciendo la producción de glucógeno (polímero de mucho mayor peso molecular) para su almacenamiento intracitoplasmático sin problemas osmóticos. Si se sobrepasa la capacidad de almacenamiento de glucógeno por el hígado (no mayor del 5-6 % de su peso) y por los músculos, un eventual exceso de glucosa-6-fosfato en sus células puede promover la glucólisis, llevando a la producción de acetil-coenzima A, ácidos grasos y triglicéridos que, en ausencia de consumo, y también por acción de la insulina (por estímulo de la lipoproteín-lipasa de los adipositos, que capta ácidos grasos, y la provisión del glicerol-fosfato para la síntesis de triglicéridos), tienden a ser acumulados en el tejido adiposo (6). La ausencia de insulina dificulta la utilización y el almacenamiento de glucosa, y promueve la utilización del tejido adiposo y el consiguiente vuelco de ácidos grasos libres de muy fácil utilización a la circulación (ciclo “glucosa-ácidos grasos libres”). El glucagón es otra hormona peptídica que estimula la producción de glucosa por el hígado mediante el desdoblamiento del glucógeno (glucogenólisis) y la utilización de moléculas de aminoácidos (neoglucogénesis), volcando el producto a al circulación y contribuyendo de esta manera a incrementar la glicemia. La secreción de glucagón es también estimulada por la concentración sanguínea de alanina (aminoácido liberado por los músculos bajo la acción de las hormonas catabólicas (cortisol) y de las enzimas proteolíticas durante el ejercicio). La adrenalina (epinefrina) y al noradrenalina (norepinefrina), hormonas conocidas en conjunto como catecolaminas, son moléculas amínicas pequeñas producidas en proporción de 4 a 1 por el sistema adrenérgico. Este está constituido por la médula suprarrenal y las glándulas nerviosas de las neuronas efectoras del sistema nervioso simpático (estructura que prepara al organismo para las situaciones en las que las únicas alternativas son la agresión o la huida), que estimulan la secreción de catecolaminas por la médula suprarrenal y producen, además, cantidades importantes de noradrenalina por su propia cuenta como mediador químico sináptico, esta secreción obedece a estímulos de otro tipo, de modo que la concentración sanguínea de noradrenalina es normalmente 5 veces superior a la de adrenalina (3). Las catecolaminas son reconocidas por dos tipos de receptores: los denominados alfa (afectados por las dos hormonas) y los beta adrenérgicos (afectados sólo por la adrenalina), de los cuales una clase particular, los conocidos como “beta-2”, tienen mayor importancia para el ejercicio físico, porque gobiernan la producción de calor y la glucogenólisis, especialmente en los músculos, aunque también, en menor medida, en el hígado. La adrenalina (y, en menor grado, también la noradrenalina) intervienen, pues, en la regulación de la glicemia mediante un proceso que contempla la integración de los sistemas nervioso y endocrino, tendiendo a aumentarla cada vez que se reduce, e incrementando la producción de glucosa mediante el estímulo de la degradación del glucógeno muscular y hepático.
La somatotrofina, u hormona del crecimiento (“growth hormona”, GH o STH) es un polipéptido secretado por la adenohipófisis en respuesta a la acción de otro polipéptido: el factor liberador de GH (“GH-releasing factor”, GHRF) producido por el vecino hipotálamo, en respuesta a la hipoglucemia o a estímulos neurológicos diversos (otro ejemplo de integración neuroendocrina) y transportado a la hipófisis por un delicado sistema local de circulación portal. La GH estimula la síntesis de proteínas y la lipólisis, reduciendo indirectamente el consumo de glucosa por aumentar la oferta de combustible alternativo (efecto estrictamente “anti-insulínico”), durante el ayuno y/o el ejercicio físico.
El cortisol (hormona principal) y la cortisona, denominados en conjunto “glucocorticoides” por efectos sobre el metabolismo hidrocarbonato, son producidos por la capa fasiculada de la corteza suprarrenal en respuesta a:
1) A su hormona “madre” hipofisaria, la adenocorticotrofina (“adrenocortico-tropic hormona”, ACTH), a su vez estimulada por el factor liberador de ACTH (“corticotrofin releasing factor”, CRF) producido por el hipotálamo en respuesta a la caída de la concentración sanguínea de cortisol (mecanismo regulatorio retroalimentado intrínseco del eje hipotálamo-hipofiso-suprarrenal);
2) A la hipoglicemia; o 3) a estímulos de orden neurológico (“stress” emotivo), con lo que se configura un nuevo enlace funcional neuroendocrino. Los glucocorticoides actúan mediante mecanismos distintos de los que involucran la producción de AMP cíclico, por los cuales se promueve el catabolismo proteico y la consecuente provisión de aminoácidos para la resíntesis hepática de glucosa (gluconeogénesis) a cargo del glucagón. Las hormonas tiroideas (de las cuales la más importante cualitativamente es la tetrayodotironina o tiroxina, T4 y la más activa es la triyodotironina, T3) son aminoácidos iodados producidos en respuesta a la hormona “madre” tirotrofina (TSH) hipofisaria, a su vez estimulada por el factor hipotalámico liberador de TSH (el llamado TRF), cuya secreción se activa cada vez que baja la concentración sanguínea de T3-T4. Las hormonas tiroideas actúan sobre casi todas las células del cuerpo, activando en general su metabolismo aeróbico, su síntesis proteica, su lipólisis y su glucogenólisis, de modo que potencian los efectos de las otras hormonas que utilizan el mecanismo del AMP cíclico en el mismo sentido (efectos “permisivo”). Además, estas hormonas tienden a aumentar la glicemia en los períodos post-prandiales, favoreciendo la absorción intestinal de glucosa.
La Figura 4 esquematiza, de manera sobresimplificada a los fines didácticos, la integración de las diversas interrelaciones funcionales que configuran el complicado sistema regulatorio retroalimentado que gobierna la glicemia, tanto en reposo como durante el ejercicio (7).
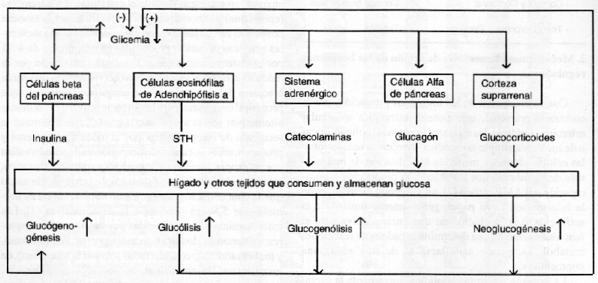
Figura 4
3. Funcionamiento del sistema regulatorio durante el ejercicio
Aún durante el ejercicio de intensidad moderada, los requerimientos musculares de glucosa como combustible de primera elección para la resíntesis de ATP-CP ocasionan una tendencia a la hipoglicemia, que en principio es neutralizada por la capacidad del hígado y en menos medida, del riñón, para liberar glucosa a la circulación; si la duración y/o la intensidad del ejercicio lo determinan, la captación de glucosa puede ser suficiente como para hacer descender realmente la glicemia y, con ella, la secreción de insulina por el páncreas. Esta circunstancia (a) reduce la captación de glucosa por todos los tejidos inactivos, y (b) aumenta la lipólisis y la provisión y utilización alternativa de ácidos grasos libres por todo el organismo, incluyendo los músculos. Este mecanismo permite un importante ahorro de glucosa para proveer al trabajo de los músculos y del cerebro. Como el consumo de glucosa por parte del cerebro es independiente de la insulina, el ajuste permite preservar prioritariamente su funcionamiento por sobre del de cualquier otra estructura, inclusive los músculos. El catabolismo proteico muscular, estimulado por el aumento de la secreción de glucocorticoides promovido por el ejercicio, resulta en una liberación de alanina a la circulación que junto con la relativa hipoglicemia concomitante, constituyen estímulos importantes para la secreción pancreática de glucagón. Esta hormona contribuye a mantener la glicemia estimulando la neoglucogénesis hepática a partir de esas moléculas de alanina, y además, incentivando la glucogenólisis. Por este mecanismo se pueden aportar como mínimo 100 gr. Diarios de glucosa a la circulación (unas 400 kcals) durante la actividad normal y en ayunas, correspondientes a las necesidades obligadas del sistema nervioso central (8).
La secreción de catecolaminas sólo se eleva si el ejercicio es más que moderado, o muy prolongado; pero si su intensidad alcanza un 50-70 % del VO2 máx. estas hormonas aumentan mucho y muy rápidamente, principalmente por aporte del sistema nervioso, el cual puede también contribuir, por estimulación neuropsíquica en la pre-competencias, al logro de niveles de catecolaminas algo mayores de lo normal al principio del ejercicio. La respuesta muscular al aumento de adrenalina, que involucra cambios en la concentración de calcio iónico intracelular, consiste en una estimulación de la glucogenogénesis, fenómeno que también ocurre en el hígado si el ejercicio es muy intenso. El resultado es una mejor oferta de glucosa para retrasar el agotamiento del sistema energético anaeróbico láctico, que puede ser beneficiosamente prematuro si el ejercicio es suficientemente intenso. La adrenalina estimula además a la lipasa hormona-sensible del tejido adiposo (6), aumentando el paso de ácidos grasos libres a la circulación y mejorando también, de esta forma, la capacidad de defender las reservas de glucosa mediante un aumento de la oferta de combustible alternativo a los músculos.
La secreción de somatotrofina es estimulada durante el ejercicio por mecanismos aún poco conocidos, entre los cuales ni la hipoglucemia, ni el aumento del lactato o la alanina sanguíneos, ni el descenso del ph de la sangre, ni el calentamiento del cuerpo, parecen figurar en forma significativamente importante, pensándose por el momento que ciertos factores neurológicos podrían ser los responsables (8). El principal efecto de la somatotrofina es aumentar la lipólisis por estímulo de la lipasa del tejido adiposo, incrementando la oferta de ácidos grasos libres como combustibles muscular alternativo de la glucosa, reduciendo de esta manera el consumo de está última en forma indirecta. A diferencia de las catecolaminas, con las cuales guarda una relación de efectos en cierto modo complementaria, la somatotrofina ejerce esta acción en forma relativamente tardía, de modo que la provisión de ácidos grasos libres durante el ejercicio estaría activada al principio por las catecolaminas, y a partir de un cierto tiempo (estimado en más de 15 minutos) por la GH o STH. Durante el ejercicio intenso y prolongado, la hipoglucemia y el “stress” pueden contribuir a incrementar la secreción de CRH hipotalámica y de ACTH hipofisaria y, en consecuencia, también al de glucocorticoides, hormonas que contribuyen a la bioenergética muscular estimulando las reacciones proteolíticas para aportar aminoácidos como combustibles alternativos de la glucosa. El recurso no es el ideal, dado que las proteínas son constituyentes plásticos específicos muy importantes en cualquier estructura viviente, incluyendo los músculos, pero su puesta en marcha sólo tienen lugar cuando la intensidad del ejercicio lo requiere, so pena de exposición a males metabólicos mayores, derivados de la importancia de una adecuada y rápida reposición de los niveles de glicemia post-ejercicio para el funcionamiento adecuado del sistema nervioso. La liberación de aminoácidos (principalmente alanina) resultante del catabolismo proteico contribuye al aporte de nueva glucosa a favor del llamado “ciclo de la alanina-glucosa” (9), mediante el cual cada molécula de alanina producida es transportada por la circulación al hígado, tejido en el que la glicólisis no está tan exageradamente estimulada como en los músculos durante el ejercicio, y por tanto tampoco está forzada la producción de lactato a expensas del piruvato. La transformación de alanina en piruvato por parte del hígado, en consecuencia, provee sustrato para la gluconeogénesis en el sitio adecuado, estimulada por el aumento concomitante del glucagón ocasionado por el mismo ejercicio. Esto posibilita el aporte de glucosa al músculo mediante la circulación en momentos especialmente críticos para el aporte energético al sistema anaeróbico lactácido de la contracción, o en el peligroso período de reposición obligada del glucógeno y de la glicemia post-ejercicio.
Las hormonas tiroideas sólo actúan indirectamente en el mecanismo analizado, promoviendo las acciones de las demás hormonas mediante un estímulo general de los procesos energéticos aeróbicos, con un aumento relativo de la síntesis proteica y del consumo de lípidos y de glucógeno.
4. Efectos del entrenamiento sobre el funcionamiento del sistema regulatorio
Podría pensarse, a priori, que el efecto general del entrenamiento sobre la secreción de las hormonas reguladoras del metabolismo glúcido debiera ser el de aumentar su producción, es decir, “incrementar la capacidad de respuesta” de las glándulas productoras ante los estímulos naturales descriptos.
Sin embrago, no resulta difícil comprender que (a) si lo que realmente se entrena es la capacidad de los sistemas enzimáticos para catalizar las correspondientes reacciones del metabolismo; y (b) si de la eficiencia de los mecanismos implicados resultara precisamente una menor variación de las concentraciones de los metabolitos cuyos valores se pretende controlar, la respuesta a esperar por parte de las glándulas productoras de hormonas reguladoras debe ser realmente menos marcada, luego que antes del entrenamiento.
Esto equivale a decir que, junto con el logro de un trabajo metabólico “más cómodo” bioquímicamente hablando, el entrenado consigue también un menor nivel de exigencia para sus sistemas regulatorios retroalimentados. Efectivamente, las respuestas (positivas o negativas) al ejercicio por parte de las glándulas productoras de insulina, glucagón, catecolaminas, somatotrofina, glucocorticoides y hormonas tiroideas son suavizadas por el entrenamiento, consecuentemente con la manifestación de un grado de control metabólico, por lo menos igual que el que existía antes de comenzar el plan. Este fenómeno, aparentemente paradójico, debe hacer reflexionar al entrenador, ya que constituye una evidencia plena de que la existencia de los sistemas regulatorios retroalimentados negativos representan un obstáculo, antes que un elemento favorecedor, para la velocidad que espera imprimir a los cambios de conductas (es decir, a las “derrotas” a infringir a los sistemas precisamente encargados de neutralizar toda clase de cambios).
Justamente, el logro de un menor nivel de actividad para un determinado sistema regulatorio retroalimentado como producto de un entrenamiento debe constituir un elemento de juicio favorable para la marcha del mismo, por cuanto expresa la natural tendencia del trabajo de sus estructuras participantes a reducirse como resultado de un accionar más coordinado de todos los mecanismos implicados, y de una consecuente mayor facilidad para arribar a los estados estacionarios tan deseables dentro de límites de variación aceptables, sea durante el reposo, en el transcurso del ejercicio, o cuando tiene lugar la recuperación.
Por el contrario, la producción de respuestas más amplias por parte del sistema regulatorio debe interpretarse como indicador desfavorable de la marcha de un entrenamiento, debido a que los organismos priorizan la persistencia de su capacidad de mantener constantes su composición y su propiedad de reponer los elementos significativos para la organización de sus unidades constitutivas (autopoyesis (2)) frente a la posibilidad de manifestar cualquier clase de cambio. Si la pretensión de forzar un cambio produce una situación de exigencia demasiado grande para los sistemas regulatorios, el sujeto, lejos de entrenarse, se está enfermando.
Si, en cambio, la misma pretensión se va satisfaciendo mientras se va logrando cada vez más “reposado” grado de los sistemas regulatorios, puede interpretarse que el grado de entrenamiento alcanzado, cualquiera fuera su magnitud, está proporcionando también un cambio favorable en los recursos homeostáticos del entrenado; es decir, esta mejorando su estado de salud.
Ningún entrenador puede autoconsiderarse eximido de la obligación de recordar que:
- El gobierno de la homeostasis es esencialmente democrático.
- Entrenar es cambiar.
- Los sistemas regulatorios se oponen democráticamente a los cambios, aunque sus resultados sean previsiblemente favorables.
- En toda democracia, si los opositores reducen el tono de voz, es porque el gobierno es bueno.
Referencias
1. Ferretti J.L (1989). Nociones fundamentales sobre Bioenergética Humana. ABCD Nro. 1: 4-9, (reedición)
2. Ferretti J.L (1989). ¿En que consiste la vida?. ABCD Nro. 2: 2-8, (Reedición)
3. Ferretti J.L (1989). El razonamiento científico. ABCD Nro. 4: 12-20, (Reedición)
4. Ferretti J.L (1990). Sistema regulatorio biológico. Parte I. ABCD Nro. 4: 2-6, (Reedición)
5. Ferretti J.L (1990). Sistema regulatorio biológico. Parte II. ABCD Nro. 4: 2-6, (Reedición)
6. Ferretti J.L (1989). Papel de los lípidos en el metabolismo energético. ABCD Nro. 3: 10-16, (Reedición)
7. Marks V, Rose F.C (1967). Hipoglucemia (Hypoglucemia, Blackwell Scientific Publications, Oxford, 1967). Toray, Barcelos, 1a Rd
8. Brooks G. A (1984). Exercise Physiology: Human Bioenergetics and its Aplications (Cap. 9). Brooks G. A., Fahey T. D. (Eds.) MacMillan, New York
9. Ferretti J.L (1990). Dietas hiperproteícas y anabolismo. Mitos y realidades científicas. ABCD Nro. 4: 29-33, (Reedición)
Cita Original
Ferretti J. L.Hormonas y Ejercicio. Actualización en Ciencias del Deporte; Vol. 5, Nro. 14, 4-12, 1997.
Cita en PubliCE
José L Ferretti (1997). Hormonas y Ejercicio. .https://g-se.com/hormonas-y-ejercicio-126-sa-j57cfb27105f2e